Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Research Article - (2013) Volume 2, Issue 4
Background: A major cellular change in dopaminergic neurons leading to Parkinsonism is the alteration of microtubule proteins that causes accumulation of tau protein, α-syn and β-amyloid plaque in the cells. In this study we investigate the role of Vitamin D3 in relieving the symptoms of Parkinsonism as it is capable of stimulating polymerization of microtubules. The Microtubules (MT) system in the fish scale melanocytes has been modeled for the dopaminergic neurons of the Substantia Nigra (SN). These cells are capable of forming cellular processes similar to what is seen in the dopaminergic neurons; in this study, we investigate the protective effect of Vitamin D3 Receptor Agonist (VDRA) and N-Methyl-D-Aspartate Receptor (NMDA R) inhibition in process formation, synaptic denervation and melanin loss in fish scale melanocytes modeled as pigmented adrenergic cells.
Method: The Tilapia scale was isolated and sub cultured in Ringer’s solution following which the cells were prepared for imaging. We incubated the cells with VDRA, Ketamine and a combination of Ketamine and VDRA in separate set ups for 60 minutes. Using brightfield imaging techniques, the cells were viewed during the incubation period and recorded using a Cameroscope connected to a computer interface.
Results/Conclusion: The cells incubated with VDRA and NMDA R inhibitor, showed an increase in the number of process and extent of the process formation; the increased number of process is an indication of a rapid rate of polymerization of microtubules. Also, the processes formed are combined long processes peculiar to the NMDA R1 inhibition and short processes characteristic of VDR potentiation as seen in VDRA treatment only. Most of the effects of the VDRA were restricted to process formation around the cell body; this is similar to the microtubule cytoskeletal system found in the dendritic nucleation assembly. This finding confirms the presence of VDR and its likely restriction to d cell body plus its role in facilitating short dendrite-like process formation while NMDA R is located on the processes and facilitates long process formation.
<ACSF: Artificial Cerebrospinal Fluid; VDR: Vitamin D3 Receptor; VDRA: Vitamin D3 Receptor Agonist; ATP: Adenosine Trios Phosphate; DNA: Deoxyribonucleic Acid; NMDA R: N-Methyl- D-Aspartate Receptor; PD: Parkinson Disease; SN: Substantia Nigra; MT: Microtubule; MTOC: Microtubule Organization Centre; NDD: Neurodegenerative Disease
Selective vulnerability of the dopaminergic neurons and pigment cells of the SN cannot be over emphasized in the etiology and progression of PD. Deficiency of Vitamin D3 or genetic mutations involving the VDR have been described in multi array organ malfunction especially in the SN, parathyroid gland, bone and gastrointestinal tract [1,2]. Vitamin D3 Receptors (VDR) have been observed in the Substantia Nigra (SN) [3,4]; as well as in the skin and liver of the fish [5]. Our choice of the VDR and VDRA, in this study, is to establish in part, the cellular mechanism of Vitamin D3 as a neuroprotective agent in PD especially as it involves process formation and prevention of synaptic denervation via signaling pathways that facilitates polymerization of MT. The model used for this study involved melanocytes in Ringer’s solution containing NMDA R blocker ketamine; to induce process and synapse-like structure formation in these cells in vitro. Thus co-treatment, in vitro, with ketamine and Vitamin D3 (VDRA) describes the pharmacological action of NMDA R inhibition and VDR potentiation. The mechanism of the VDRA involves calcium signaling pathways that facilitate polymerization of MT such that the αβ-tubulin sub-units assemble and convey melanosomes and other vesicles to the periphery of the cell using the kinesin motor assembly [6]. This is believed to be complimentary to the inhibition of NMDA R; prevention of glutamate toxicity [7], as such movement of vesicles, away from the nucleus prevents cell death by autophagy.
The VDRA binds to a cytoplasmic receptor VDR having two domains; first is the hormone binding domain and second is the DNA binding domain. A VDR-VDRA receptor complex is formed and can bind to the promoter region of genes, thus increasing the rate of gene transcription and cellular activity [8]. Several studies have also shown that VDRAs are members of neuroactive steroids capable of regulating several pathways important in neuronal development and function [9]. There is growing evidence that Vitamin D deficiency causes alteration in dopaminergic neurons in the adult brain [10]. Although the presence of the VDR have been described in the SN, there is no concrete evidence to describe the presence of this receptor in pigment cells or adrenergic cells and also no recorded evidence as to how they moderate cellular activities in the SN [11]. Other studies have shown that presence of corticosteroids like hydrocortisone did increase the gene transcription rate of VDR genes detected using RT-PCR; an act which is complementary in VDR up-regulation [12]. We are modeling the melanocytes in fish scale as the adrenergic neurons MT and pigment system [13,14]. In addition both cells are sites of action for catecholamines (epinephrine and nor epinephrine) and monoamines (Dopamine), thus we can deduce some information about signal transduction property of pigment cells endowed with special MT system for signal regulation and coordination [15].
The fish oil have also been found to be rich in Vitamin D3, as this oil has been found to be abundant in the skin of the fishes, also Vitamin D3 receptors have been found in the brain, thus suggesting the relationship between MTs, melanocytes, neurons and Vitamin D3 activities [16]. Vitamin D3 is capable of stimulating MT activity and regulating the aggregation and dispersion of pigments in the scale of fishes, in the skin of humans. Deficiency of Vitamin D3 has been found to cause a disease called ricket, characterized with melanin patches on the skin, showing the dysregulation of proteins required for dispersion of melanin in the skin [17,18]. Vitamin D3 has also been observed to have a major influence in the exchange of calcium and phosphorus in the gastro intestinal tract and the Harvesian system of the bone [19]. This study is aimed at demonstrating melanocytes as a suitable model for MT and pigmented adrenergic cells of the Substantia Nigra (SN). The role of VDR potentiation and NMDA R inhibition was investigated in fish melanocytes; especially in the aspect of cellular process formation and synaptic denervation in the modeled pigment cells.
All reagents were procured from Sigma-Aldrich, Germany. The ketamine and Vitamin D3 were procured from Kanada Pharmaceuticals, Ekiti State, Nigeria.
Investigating MTs in fish scale in vitro
Freshly harvested fish scale (Tilapia) was placed in cold freshly prepared Ringer’s solution with the following composition; Sodium chloride: 8 g, Potassium Chloride: 0.42 g, Calcium Chloride: 0.24 g and 0.20 g of Sodium bicarbonate. The solution was made up to 1,000 ml in double distilled water using the method of Singh and Lal [20].
Treatment and imaging
Vitamin D3 (VDRA) was used at a concentration of 1.5 micro/ Kg and NMDA R1 antagonist (ketamine) was used at a concentration of 25 μM. Four separate set ups were mad as follows; A: Ringers Solution, B: Ringers Solution+Ketamine, C: Ringers Solution+Vitamin D3+Ketamine, D: Ringers Solution+Vitamin D3. The process of cellular changes was recorded using a 5.1 MP Cameroscope connected to a bright field microscope (Olympus) and computer interface for live cell recording. No contrast media was added to the cultures; the cells were also unfixed. The culture was maintained by perfusion at an interval of 5 minutes and this was carried out for a maximum duration of 60 minutes for each of the separate treatment categories (A, B, C and D).
Image acquisition
The images were rendered in gray scale using Image J (from NIH). Measurement of cellular processes was done by inverting the gray scale to create a white image on a dark background and to enhance contrast. The images were analyzed on Open Office Draw Suite (JAVA) to obtain the cell process lengths in micrometers (μm).
Results
The control cells were placed in Ringer’s solution only and showed no cellular changes after 60 minutes (Figure 1A and B). The cell bodies were condensed with less prominent projections. Inhibition of the NMDA R facilitated the formation of long cellular processes resembling axons (arrows; Figure 2A-2C). The cell bodies were reduced in size compared to the control having a pyramidal appearance (arrow head). The co-incubation of the cells with VDRA and ketamine (NMDA R antagonist), caused an increase in process formation (Figure 3A- 3C). The number or processes formed increased showing a rapid rate in polymerization of microtubules; also the processes formed are combined long processes peculiar to the NMDA R inhibition (Figure 3: yellow arrows) and short processes characteristic of VDRA treatment, as seen in VDRA treatment only (Figure 3: blue arrows). Most of the effect of the VDRA was restricted to process formation around the cell body; this is similar to the microtubule cytoskeletal system found in the dendritic nucleation assembly of neurons. These findings confirm the presence of VDR and its likely restriction to the cell body, and its role in facilitating short dendrite-like process formation while NMDA R is located on the processes and facilitates long axonlike process formation. Sequel to combining VDRA treatment and NMDA R inhibition, the terminal parts of the long processes seen are characterized by blobbed ends rather than being straight endings observed in NMDA R inhibition only. Treatment with the VDRA only, facilitated the formation short cellular processes originating from the cell body (Figure 4A-4C). Morphometric analysis shows that the length of the processes observed in NMDA R inhibition is about 30 μm; while for the short processes observed in VDRA treatment are about 10 μm in length (Figures 5B). In VDRA and NMDA R inhibition cotreatment, the two forms of processes were observed, the number of process formed also increased significantly than what was observed in single treatments (Figure 5A). This further clarifies the separate roles of the two receptors as being responsible for different types of process formation in this cell model.
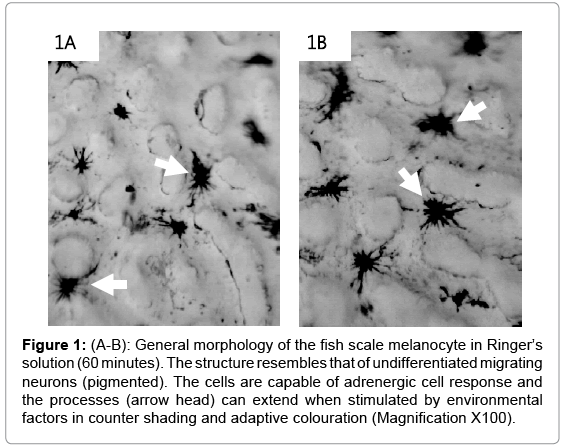
Figure 1: (A-B): General morphology of the fish scale melanocyte in Ringer’s solution (60 minutes). The structure resembles that of undifferentiated migrating neurons (pigmented). The cells are capable of adrenergic cell response and the processes (arrow head) can extend when stimulated by environmental factors in counter shading and adaptive colouration (Magnification X100).
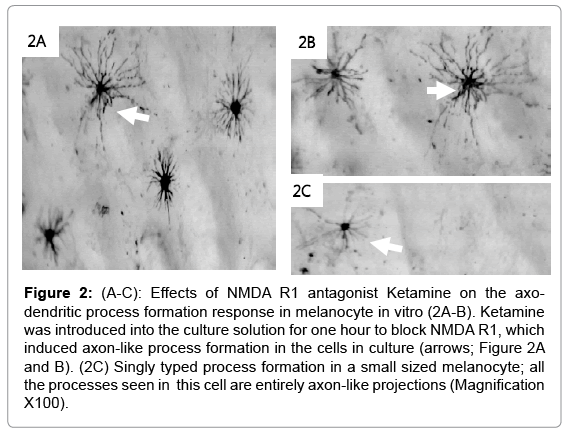
Figure 2: (A-C): Effects of NMDA R1 antagonist Ketamine on the axodendritic process formation response in melanocyte in vitro (2A-B). Ketamine was introduced into the culture solution for one hour to block NMDA R1, which induced axon-like process formation in the cells in culture (arrows; Figure 2A and B). (2C) Singly typed process formation in a small sized melanocyte; all the processes seen in this cell are entirely axon-like projections (Magnification X100).
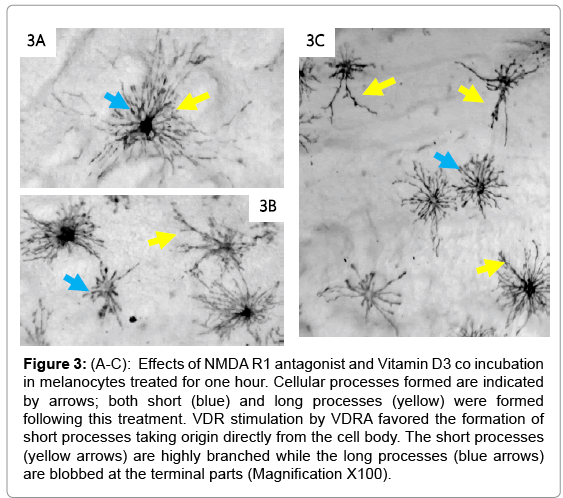
Figure 3: (A-C): Effects of NMDA R1 antagonist and Vitamin D3 co incubation in melanocytes treated for one hour. Cellular processes formed are indicated by arrows; both short (blue) and long processes (yellow) were formed following this treatment. VDR stimulation by VDRA favored the formation of short processes taking origin directly from the cell body. The short processes (yellow arrows) are highly branched while the long processes (blue arrows) are blobbed at the terminal parts (Magnification X100).
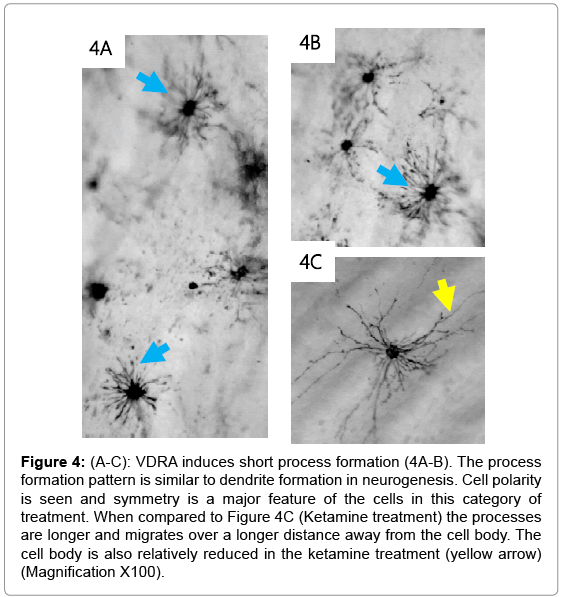
Figure 4: (A-C): VDRA induces short process formation (4A-B). The process formation pattern is similar to dendrite formation in neurogenesis. Cell polarity is seen and symmetry is a major feature of the cells in this category of treatment. When compared to Figure 4C (Ketamine treatment) the processes are longer and migrates over a longer distance away from the cell body. The cell body is also relatively reduced in the ketamine treatment (yellow arrow) (Magnification X100).

Figure 5: (A-B): Inverted gray scale cell process measurement for Control cells, Ketamine treatment, VDRA+Ketamine and VDRA Only. Increase in process length was observed in all the treatment categories. Ketamine treatment gave the longest cell process while VDRA treatment induced shorter process formation. (5B) (*P<0.001 compared to the Control) (*NMDA R1- Ketamine Treatment).
Discussion
In several experiments, nocodazole (a common agent for depolymerizing MT) has been described as a chemical substance capable of inducing cell death specifically by depolymerization of MTs and inhibition of the vesicular transport system, thus causing centralization of vesicles which in turns lead to autophagy [21]. Other studies describe the selective vulnerability of dopaminergic neurons to MT dependent degeneration [22,23]. To further elaborate on MT polymerization and depolymerization, epinephrine, norepinephrine and dopamine (adrenergic transmitters) are capable of inducing polymerization of microtubules through secondary messenger mechanism involving cyclic adenosine monophosphate (cAMP) [24,25]. In the melanocytes isolated from the Tilapia, epinephrine and cAMP drives assembly of motor proteins dyenin and kinesin from the Microtubule Organization Center (MTOC) for counter shading and adaptive coloration (aggregation versus dispersal of melanosomes) in the Tilapia [26]. We are thus anticipating that dopaminergic neurons may contain intrinsic pathways of generating epinephrine to drive a similar movement of melanin pigment in response to impulse from the association areas; similar to the findings of Nilsson and co-workers [27].
In fishes, signal is being transduced through the scales on the body surface and causes epinephrine dependent movement of melanosomes form the MTOC via motor proteins (kinesin and dyenin) [28,29]. Likening dopaminergic neurons to the melanocytes, they are not just neurons but specialized sensors of positions and environment [30]. The melanin pigments are most likely regulators of signal transduction in the entire movement coordination system of the brain, such that when nerve impulse passes, the Tubulin-motor protein complex are assembled from the MTOC or move pigments along their length in these neurons, thus generating specific information as to the nature of the movement and the extent of muscle contraction required for such movement [31]. This is also evident in the fishes as the scales have been found not to only contribute to the biological adaptation of fishes in water (counter shading) but senses the environment and contribute to movement and balance [32]. The loss of melanin pigments coupled with synaptic denervation have been implicated in the etiology of PD [3]. This can be studied using this model as depolymerization of the MT will cause centralization of all melanosomes being conveyed on such microtubule tracts. Aside having aggregation of malfolded MT proteins, accumulation of vesicles will induce cell death by autophagy [33]. In a similar way, synaptic denervation is a function of depolymerization of the axo-dendritic cytoskeletal structure [34]. In this study we have used Tilapia because it is readily available, the scales and cells are large and will require little magnification for clarity of melanocytes. The protein sequence for the MT protein is exact match with that of the rats and human models.
The role of VDR and NMDA R in such process formation cannot be over elucidated. NMDA R1 have been described as a specialized glutamate receptor that facilitate cell migration, process formation and synapse formation in developing neurons while VDR is capable of generating calcium based signal transduction which can facilitate polymerization of microtubules; although the function of the NMDA R differ slightly in the developing and adult neurons [35-37].
The studies of Cui et al. [38], suggests that VDR is abundant in dopamine rich regions of the brain, also highly hydroxylase positive neurons in humans expressed VDR. Deficiency of Vitamin D or loss of VDR is usually associated with risk of several neurodevelopmental disorders to loss of antiproliferative functions, thus the absence of VDR or VitD3 deprives the neurons of needed signals. The studies of Suzuki et al. [39] also corroborated this finding that Vitamin D deficiency is a predisposing factor to increased progression of PD. The findings of this study suggest that the progression of such disease conditions is related to process formation and synaptic denervation. This study further elaborates the cellular mechanism involved in the disease progression. This study however shows that inhibition of the NMDA R did facilitate the formation of long axon-like processes extending between two adjacent melanocytes, while the VDR stimulation caused short process formation radiating from the cell body and representing an array of bi-directional vesicular transport system. Similar to our proposition in this study, Aspengren et al. [40] suggested that Melanocytes develop from the neuronal crest and are most abundant in the dermal and epidermal layers of the skin, where the intracellular distribution of the pigment significantly influences the color of the animal. The transport of pigment is dependent on an intact cytoskeleton and motor proteins associated with cytoskeletal components. From a neuroscience perspective, other studies have also shown that axon dendrite process formation is differentially affected [36]. The control cells were placed in Ringer’s solution and showed no cellular changes (Figure 1A and B). The cell bodies are condensed with less prominent projections. Inhibition of the NMDA R facilitated the formation of long cellular processes resembling the axons (Figure 2A-2C). The cell bodies were reduced in size compared to the control having a pyramidal appearance (arrow head). Adjacent cells also maintained temporary contact similar to what is obtainable in neuronal synapse formations.
Conclusion
The co incubation of the cells with VDRA and NMDA R inhibitor, Showed an increase in process formation. The number or processes formed increased showing a rapid rate in polymerization of microtubules; also the processes formed are combined long processes peculiar to the ketamine treatment and short processes characteristic of VDR treatment as seen in VDRA treatment only. Most of the effects of the VDRA were restricted to process formation around the cell body; this is similar to the microtubule cytoskeletal system found in the dendritic nucleation assembly. This finding confirms the presence of VDR and its likely restriction to d cell body plus its role in facilitating short dendrite like process formation while NMDA R is located on the processes and facilitates long process formation; similar to the findings of Beckerman et al. [37].
Parkinson’s disease (and other associated movement disorders) is a common condition that is triggered by chemical, environmental, genetic and neurotrophic factors in which dopaminergic neurons are lost and melanin pigment is reduced in the SN. If not treated, it can progress rapidly into movement disorders and other cognitive dysfunctions. Since the initial description of this disease and other NDD’s, efforts have been directed towards understanding the molecular mechanism of pigment loss and cell death in the SN. The cause of PD has been broadly described as unclear; in addition loss of adrenergic cells have also been described in the progression of the diseases condition which is characterized by shortening of neuronal projection as a form of synaptic denervation. However, to understand the process involved in this synaptic denervation of adrenergic pigmented neurons, the molecular mechanism needs to be studied at cell and protein level.
In this study, we have first described the mechanism involved in neuronal loss in the SN and the general structure of the fish scale melanocyte. Using bright field imaging techniques, live cell imaging was conducted in vitro to show the various changes observed in the melanocytes upon manipulation of the NMDA R and the VDR. The outcome shows that the fish scale melanocyte contains NMDA R on its membrane just like the human neuronal cells, although it is much more localized on the axon-like processes; while VDR is localized on the cell body and short dendrite-like processes. Upon inhibition of this receptor (using ketamine; a human NMDA R antagonist), the cell projections forms wide array networks of cellular processes following a similar pattern to what is observed in neuronal axon-dendrite formation. Glutamate treatment (NMDA R potentiation) also caused formation of cellular projections but not as extensive as that seen in NMDA R inhibition. These findings therefore creates a premise for the study of pigmented neuronal cells in vitro as this cell type (fish scale melanocyte) expresses NMDA R and the role of this receptor in cellular process formation has also been seen to be similar to the pattern observed in neuronal axon-dendrite formation. Inverted gray scale image analysis shows that this cell upon inhibition of NMDA R shows temporary connections between the formed processes, an association similar to the synapse that has been observed in the human neurons. VDR stimulation facilitated more of short process formation radiating directly from the cell body suggesting its role in cellular process formation similar to that of the dendritic nucleation assembly in neurons.
We acknowledge the resources shared and provided by the Alzheimer’s Drug Discovery Foundation (ADDF). The Directorate of Research, Afe Babalola University and the Laboratory Staff of the Department of Biological Sciences. Also we acknowledge the contributions of Elizabeth Peters of the College of Medicine for assisting in the revision of this work.