Journal of Oceanography and Marine Research
Open Access
ISSN: 2572-3103
ISSN: 2572-3103
Research Article - (2013) Volume 1, Issue 1
Palytoxin (PLTX) is a complex marine toxin synthesized by the soft coral Palythoa toxica and by species of the benthic dinoflagellate Ostreopsis spp. The toxin binds to the active Na, K-ATPase pump in the cellular membrane. This interaction changes the protein conformation and produces a non-specific cation channel. Using the Fluorescent Polarization (FP) technique to quantify the PLTX concentration, the production of PLTX-like products was measured in several cultures of Ostreopsis ovata and Ostreosis siamensis. The cultures were grown under different conditions to study the optimal parameters to grow and to produce toxins. Serious difficulties were found to quantify the number of cells at the end of the exponential phase. In order to avoid any toxin loss, the weight of the pellet obtained after careful filtration was used as reliable parameter to calculate the growth of the cultures. Also, this parameter was used to refer the results of toxin concentration. In addition the toxicity of the cultures was measured by mouse bioassay. In these conditions, the optimal parameters to grow these strains are 24°C, 37‰ of salinity and 16:8 h light-dark photoperiod. Within these parameters high amounts of PLTX-like compounds with different toxin profiles were obtained.
<Keywords: Palytoxin; Na,K-ATPase; Ouabain; Ostreopsis spp; fluorescent polarization; Cleaning method
Palytoxin (PLTX) is a toxic compound initially isolated from marine soft corals of the genus Palythoa [1] but also produced by tropical benthic dinoflagellates from Ostreopsis genus [2]. This toxin has been involved in human poisoning after the ingestion of food contaminated, but also skin irritations and respiratory effects by contact with contaminated spray seawater have been reported [3].
The PLTX-group of toxins includes complex polyhydroxylated compounds with a lipophilic and a hydrophilic area. Different molecules related with the PLYX structure (Figure 1) have been described: palytoxin-b, homopalytoxin, bishomopalytoxin, neopalytoxin, deoxypalytoxin, 42-hydroxypalytoxin, ostreocin-d, ovatoxin-a, -b,-c,-d, -e and -f and mascarenotoxin -a, -b and -c. However only the chemical structures of PLTX, ostreocin-d, ovatoxin-a and 42-hydroxypalytoxin have been characterized [4-11]. PLTX is a large molecule, with a molecular weight of 2659-2680 daltons depending on the origin [12,13].
Figure 1: Chemical structure of Palytoxin.
The target of PLTX is the Na+,K+-ATPase pump from the plasma membrane. This pump exchanges three Na+ ions from the cytosol to the extracellular space for two K+ ions from the outside of the cell into the cytoplasm, expending one ATP molecule [14]. The toxin produces a free flow of ions, and as consequence the cationic permeability of the cells is altered. This imbalance in excitable cells causes symptoms like scratching, jumping, paralysis of hind limbs, respiratory distress, cyanosis also brings nausea, tiredness, diarrhoea and vomiting followed by dizziness, in animals, but also intestinal symptoms, muscle spasms, breathing difficulties followed in some cases by dead in humans [15-17]. This type of human poisoning is called palytoxicosis or clupeotoxicosis [18].
Ostreopsis spp. is unicellular algae. The habitat and the growth of these dinoflagellates are very important in toxin production [5]. Temperate tropical coastal waters were the original localization of these species [19], however these blooms have been described in other localizations far away from tropical waters, as the Thyrrenian and Adriatic Seas [19,20] Spain [21,22] Greece [23] and more recently in Portugal [24]. Therefore the general rise of water temperatures may be modifying the Ostreopsis spp. world distribution, and therefore these dinoflagellates can appear in new places where they never have been seen before, becoming PLTXs compounds a serious public health problem.
Ostreopsis spp. grows better in calm, shallow, warm waters and it seems that high salinity concentration increases their development [25-27]. But it is not clear the importance of the salinity and nutrients in their blooms [26]. Therefore it is essential to know the optimal conditions for the growth of these cells and the link with toxin production. Therefore, in this context, the aim of this work is to estimate the production of PLTX-like compounds in different cultures of Ostreopsis spp. growing in several biological conditions by using a fluorescence polarization (FP) technique recently developed [28].
Chemicals
PLTX was obtained from Wako Pure Chemical Industries (Neuss, Germany) and it was dissolved in MeOH:H2O (1:1). Adenosine 5-triphosphatase (Na,K-ATPase) from porcine cerebral cortex and ouabain were obtained from Sigma-Aldrich Química, S.A. (Spain); Fluoro-Spin 498 Protein Labeling Kit was obtained from EMP Biotech (Germany).
Methanol, n-hexane and n-butanol were obtained from Panreac (Spain). LC-18 SPE tubes 6 mL and 2 mL Empty PP Rev Tube Kit with silica (600mg) were obtained from Sigma-Aldrich Química, S.A. (Spain). Ultrafree-CL filters (0.45 μm pore diameter and low-binding regenerated cellulose as membrane) were purchase from Millipore (Spain).
The composition of PBS is: NaCl 137 mM; Na2HPO4 8,2 mM; KH2PO4 1,5 mM; KCl 3,2 mM y MgCl2 1 mM.
Dionized water (Milli-Q) was used for all experiments. PBS chemicals were obtained from Panreac Quimica S.A.U. (Spain), except KH2PO4 which was purchased from Merk (Spain). Saline solution was obtained from Laboratorio Grifols S. A. (Spain) and Tween® Monostearate (Tween 60) from Sigma-Aldrich Química, S.A. (Spain).
Protein labeling and Fluorescence polarization measurements
To label Na,K-ATPase, a reactive succinimidyl-ester of carboxifluorescein was used. This molecule forms a stable covalent linkage between the protein and the dye. This conjugation has fluorescent excitation and fluorescence emission maxima at 498 nm and 522 nm respectively. The Fluoro Spin 498 Protein labeling kit contains all of the chemicals and tools needed during the labeling and purification processes.
The protein was mixed with a sodium bicarbonate solution, then the dye was added, and the mixture reacted for 1 h at room temperature while protected from light. Afterward, a hydroxylamine solution was added to stop the labeling and incubated for 30 min. The final conjugate (Na,K-ATPase - F) was purified (separated from the non attached dye) by using spin columns. The conjugates can be stored for a long time in frozen aliquots at -20°C while protected from light. With this protocol 2 mg/mL of adenosine 5-triphosphatase (Na,K-ATPase) from porcine cerebral cortex were labeled. The labeling ratio was 17 molecules of dye per molecule of protein. The final conjugate was dissolved in PBS. To select the correct dilution factor to make the experiment, six different dilutions, ranging from 1:100 to 1:7500, were tested in a final volume of 250 μL. The dilution 1:5000 was chosen. With this dilution factor the final protein label concentration in each well was 2.5 ng/mL. This concentration remained constant in all experiments.
The FP is a spectroscopic technique based on exciting a fluorescent molecule with plane-polarized light and measuring the polarization degree of the emitted light. An important application of this technique is the study of molecular interactions since the FP value of a molecule will be different if a binding with another molecule occurs [29]. In each experiment as control, the cardiac glycoside ouabain 1mM, a known Na,K-ATPase blocker, was used. This drug binds to Na,K-ATPase in the same mode than PLTX. In this way, a fall of 150 millipollarization (mP) units was used as control in each experiment for the ouabain wells.
In each well 100 μL of PBS solution, 25 μL of sample (culture extract or toxin) and 125 μL of Na,K-ATPase-F were added. Then the microplate was shaken for 10 minutes at 300 rpm at room temperature, before measuring. In the control, the quantities were 100μL of PBS solution, 25 μL of ouabain 1 mM and 125 μL of Na,K-ATPase-F. In each plate PLTX calibrations curve with concentrations of: 10nM, 25nM, 50nM, 75nM and 100nM were used to calculate the unknown concentration of samples.
The Synergy 4 instrument from Biotek was used to measure the FP. The measurements were done in black polystyrene 96-well microwell plates (Costar, Spain). The light source was a Tungsten lamp. The emission light was detected by photomultipliers. Filters and polarization filters (for FP) were used to select the correct wavelength. In this case, the filters of 485 nm for excitation and 528 nm for emission were used to measure the fluorescence intensity, and the polarization filters of the same wavelength were used to measure the FP in mP units. The sensitivity was set at 100. The FP results were calculated with the Gen 5 1.07 software.
Dinoflagellate cultures
The cultures used were: Ostreopsis ovata, (A and B), Ostreopsis siamensis, Coolia monotis and Protoceratium reticulatum. The cultures were grown in seawater enriched with L1 medium, without silicates and a salinity level of 37‰. Also some cultures were grown at 42‰ salinity. The salinity was adjusted by the addition of freshwater removing chlorine by aeration. The experiments were carried out at 19°C and 24°C and the cultures were subjected to photoperiod with day-light lamps on a 16:8 h light-dark photocycle. Cells were gently shaken manually twice a day and its growth was divided in steps, increasing the volume progressively.
300 cells/ml was always used as initial inoculum and the profile and conditions of growth for each strain were previously established. In these cultures the beginning of the stationary phase of growth was reached 15-18 days after inoculation [25]. The dinoflagellates were kept under these conditions for 30 days, then the whole culture volume was carefully filtered, 20 μm mesh, and the residue obtained was weight. Finally methanol-water (1:1) was added and the extract was sonicated to their complete homogenization and toxin release and was stored frozen until measurements were to be done.
The added volume was three times the residue weight. To count the number of cells, the cultures were carefully homogenized, 25 ml pipette shaking, and 5 ml aliquots were collected. In some experiments chemical disaggregation by adding HCl to a final concentration of 4 mM was done. Cells were fixed with Lugol´s solution and were counted by optical microscopy in a Sedgewick-Rafter chamber [30].
Cleaning steps
The cultures cleaning procedure is shown in (Scheme 1) briefly, 500 μL of the cellular extract solved in methanol-water (1:1) were centrifuged 10 minutes at 5000 rpm at 4°C. The supernatant was filtered through a 0.45 μm cellulose filter. The solution filtered was evaporated to dryness and diluted in 2 mL of deionized water and charged into SPE (Solid Phase Extraction) cartridge (LC-18). Previously the cartridge was conditioned with methanol and water. Then, after loading the sample, methanol 20% was added and the toxin was eluted. This fraction was evaporated to dryness and solved in 500 μL of MeOH-H2O (1:1). This protocol previously described was extended with a filtration step, through a 10000 NMWL filter [28,29]. The filtered extract was diluted 1:1000 to the final volume of 250 μL to check the toxin concentration by FP.
Scheme 1: Scheme of clean protocol steps for Ostreopsis spp processing.
Mouse bioassay
The lethal potency of culture samples was estimated by intraperitoneal (i.p.) injection of the extracts or PLTX standard resuspended in saline solution with 1% Tween 60 into healthy male Swiss mice (SPR), 19-23 g body weight. The toxicity was determined by the time of death of six mice, according to the standard dose-lethal time plot prepared by using several dilutions of PLTX standard. The culture extract was centrifuged at 5000 rpm 10 minutes at 4°C, and the supernatant was diluted at least 1:100 in the saline solution with 1% Tween 60. Therefore less than 0.5% of methanol was injected. 1 mL of this preparation was administrated to mice. Controls of 0.5% methanol and non-toxic strains were done and no effects were observed. Mice were observed during 24 hours, the symptoms were described and also the death time.
Data analysis
All experiments were carried out at least three times using duplicates. Data were normalized and results were expressed as mean ± SEM. Results were analyzed using the Student t test. A probability level of 0.05 or less was used for statistical significance.
The aim of this work was to study the effect of different biological conditions in growth and toxin production of several strains of Ostreopsis ovata and Ostreopsis siamensis. Initially Ostreopsis spp. growth at 19°C and 24°C was studied. All cultures were grown in a total volume of 800 mL until the end of stationary phase, 30 days of growth. Then, the cultures were gently homogenated and filtered and the residues obtained were weighed and re-suspended in methanolwater. The cells were counted before filtration. With these data the growth could be represented as the number of the cells at the end of the exponential phase and also as the weight of the pellet after the whole culture volume was filtrated. Since Ostreopsis spp. are growing in lumps and glued to the edge of the container involved in a mucous substance it is very difficult to break the lumps and to do a reliable and representative counting. O. ovata A growth at 19°C and 24°C is represented in (Figure 2).
Figure 2: Effect of temperature on Ostreopsis ovata A growth. (A) Number of cells in a sample obtained after 30 days of growth. (B) Weight of pellet after filtration of 30 days cultures. Mean ± SEM of three experiments. (*) Significant differences with respect to 19ºC.
After 30 days of culture a total of 2.89 ± 0.057×106 cells at 19°C and 1.28 ± 0.4×106 cells at 24°C were obtained, (Figure 2A). Therefore, from this a significantly decrease of cells number at 24°C could be concluded. However, when the weight is represented, a significant increase of cell weight at 24°C respect to 19°C is showed, 1.76 ± 0.03 g versus 2.8 ± 0.52 g, (Figure 2B). Therefore, the results seem to be opposite taking into account the weight or the cell number. In the case of O. ovata B strain (Figure 3) the number of cells also decreases from 2.38 ± 0.1×106 cells at 19°C to 2.14 ± 0.13×106 cells at 24°C. Attending to the weight of the pellet, there is a significant increase in grams of the pellet, from 1.63 ± 0.1 g at 19°C to 4±0.36 g at 24°C. When O. siamensis was studied the results were equal, either representing cell number or the pellet weight (Figure 4); the cell number is significantly higher at 24°C, 1.5 ± 0.3×106 cells, than at 19°C, 0.28 ± 0.08×106, (Figure 4A), and the cells weight was also significantly increased from 1.07 ± 0.12 g at 19°C to 2.3 ± 0.14 g at 24°C, (Figure 4B). Therefore, in general and taking into account pellet weight, 24°C is the optimal temperature to grow Ostreopsis spp. cultures.
Figure 3: Effect of temperature on Ostreopsis ovata B growth. (A) Number of cells in a sample obtained after 30 days of growth. (B) Weight of pellet after filtration of 30 days cultures. Mean ± SEM of three experiments. (*) Significant differences with respect to 19ºC.
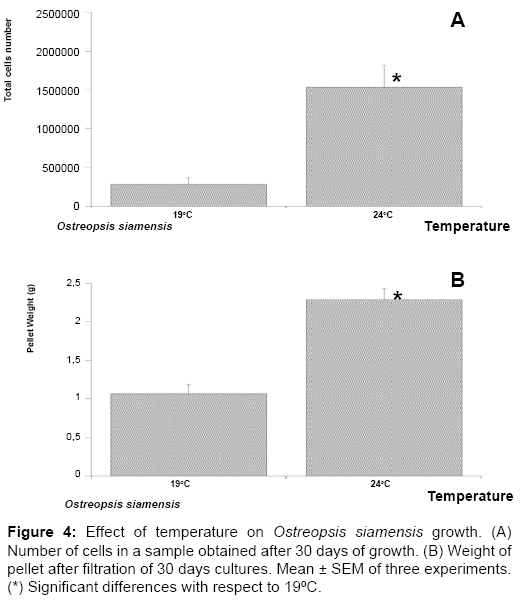
Figure 4: Effect of temperature on Ostreopsis siamensis growth. (A) Number of cells in a sample obtained after 30 days of growth. (B) Weight of pellet after filtration of 30 days cultures. Mean ± SEM of three experiments. (*) Significant differences with respect to 19ºC.
To check if the increase in pellet weight is only due to the mucus presence or to the increase in cell number the counting was repeated after cellular chemical disaggregation by adding HCl 4 mm final concentration [25,31]. Several O. ovata a cultures were grown in a total volume of 200 mL for 30 days at 19°C and 24°C. Then in some flasks HCl was added and the cell number was counted. As (Figure 5) shows, the number of cells counted with and without HCl is different at any temperature checked, however after HCl addition the number of cells at 24°C was significantly higher and therefore the increase in pellet weight match with an increase in cells number. It is important to bear in mind that HCl addition decreases seawater pH from 8.3 to 3 and cell viability after 15 minutes was 25% lower, in addition motionless cells and several debris were observed. Therefore, the following studies about toxin concentration were always referred to pellet weight since it is a reliable parameter and the cells integrity is an important issue in order to avoid any toxin leakage to the medium in the next experiments.
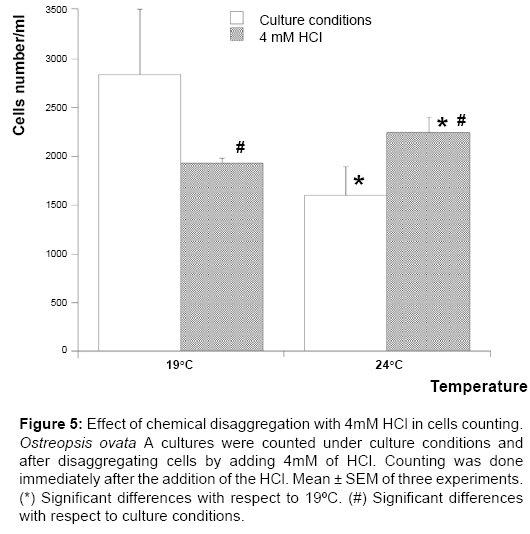
Figure 5: Effect of chemical disaggregation with 4mM HCl in cells counting. Ostreopsis ovata A cultures were counted under culture conditions and after disaggregating cells by adding 4mM of HCl. Counting was done immediately after the addition of the HCl. Mean ± SEM of three experiments. (*) Significant differences with respect to 19ºC. (#) Significant differences with respect to culture conditions.
The next point was to check the PLTX-like compounds production in these conditions. The method to quantify the PLTX is based on the FP methodology. In several studies, the FP technology was used to quantify toxins [29,32,33] and recently it was applied to develop a sensitive PLTX detection method [28]. In general, the FP units of a Na,K-ATPase-F conjugate decreased in the presence of PLTX and/or PLTX-like compounds with the same mechanism of action.
By using a calibration curve with FP units versus PLTX concentration, the amount of PLTX-equivalent in a sample can be estimated (Figure 6). To check the toxin concentration in the dinoflagellates cultures, several cleaning steps to avoid a matrix effect were done, then Ostreopsis spp. cultures and two non-PLTX producer cultures were tested as negative control, Coolia monotis and Protoceratium reticulatum. These two cultures were used to check the matrix interference and the recovery of the cleaning procedure. As (Table 1) shows, neither Coolia monotis nor Protoceratium reticulatum produce any change in Na,K-ATPase-F conjugate FP units. However, when Ostreopsis spp. cultures were analyzed, different PLTX-equivalent amounts were obtained, 113.41 ± 32.7 μM PLTX-equivalent for O. ovata A strain, 67.65 ± 12.8 μM PLTXequivalent for O. ovata B strain and 20.2 ± 8.4 μM PLTX-equivalent for O. siamensis. These strains were also analyzed by mousse bioassay and were found positives after mousse injection.
| Species | PLTX equivalent concentration (μM) |
|---|---|
| Protoceratium reticulatum | 0 |
| Coolia monotis | 0 |
| Ostreopsis ovata A | 113.41 ± 32.7 (positive mouse bioassay) |
| Ostreopsis ovata B | 67.65 ± 12.8 (positive mouse bioassay) |
| Ostreopsis siamensis | 20.2 ± 8.4 (positive mouse bioassay) |
Table 1: PLTX-like compounds concentration in different cultures extracts.
Figure 6: Effect of PLTX on FP units of Na,K-ATPase-F conjugate. Different concentrations of PLTX were added over a constant amount of fluorescent Na,K-ATPase-F conjugate. FP was measured after 10-min incubation. Mean ± SEM of three experiments.
Then the PLTX-like compounds production in different culture conditions was checked. First, the effect of salinity was studied (Figure 7); the rest of the parameters were 24°C and 16:8 h light-dark photoperiod. In these conditions, 526.4 ± 24.3 μg PLTX-equivalent/g pellet were produced by the O. ovata A strain at 37‰ of salinity, while at 42‰ the amount was significantly decreased to 265.45 ± 44.8 μg PLTX-equivalent/g pellet. For the O. ovata B strain, the production was 351.5 ± 129.4 μg PLTX-equivalent/g pellet at 37‰ and 336.7 ± 92.7 μg PLTX-equivalent/g pellet at 42‰. On the other hand, high mortality was observed when O. siamensis was grown in a 42‰ salinity medium. Therefore, 37‰ salinity was chosen to keep Ostreopsis spp. cultures. The second parameter studied was the volume of growth. The cultures were grown in a total volume of 1800 mL or 800 mL, both starting from the same initial inoculums, however no significant differences were found even when lower pellet weighs were obtained with 1800 mL (data not shown).
Figure 7: Toxin production of cultures of O. ovata A and B, growing at 37‰ and 42‰ of salinity and 24ºC. Mean ± SEM of three experiments. (*) Significant differences with respect to 37‰ of O. ovata A strain.
Then, the influence of temperature in PLTX-like compounds production was studied. The cultures were grown at 19°C and at 24°C and the rest of parameters were 37‰ salinity and 16:8 h light-dark photoperiod. As (Figure 8) shows, in all cases the toxin production is higher at 24º C. O. ovata A produces 216.75 ± 151.4 μg PLTXequivalent/ g pellet at 19°C and 768.6 ± 115.5 μg PLTX-equivalent /g pellet at 24°C. O. ovata B produces 415.25 ± 24.8 μg PLTX/g pellet at 19°C and 868.26 ± 76.5 μg PLTX-equivalent/g pellet at 24°C. In addition O. siamensis produces 45.5 ± 20.5 μg PLTX-equivalent/g pellet at 19°C and 256.6 ± 26.3 μg PLTX-equivalent/g pellet at 24°C. The production at 24°C is at least double than at 19°C and in all cases. Hence the optimal temperature for these dinoflagellates to produce PLTX is 24°C.
Figure 8: Toxin production of cultures of O. ovata A and B and O. siamensis at 19ºC and at 24ºC. Mean ± SEM of three experiments. (*) Significant differences with respect to 19ºC.
Finally, mousse bioassay was used to check the total toxicity of the Ostreopsis spp. cultures. (Table 2) shows LD50 of the PLTX standard control and also the lethality of different extracts. Several concentrations of PLTX standard, from 2000 to 500 ng/kg mouse weight, diluted in 1 mL of the saline solution with 1% Tween 60 were injected to mice. From these experiments, 750 ng PLTX/Kg mice weight was calculated as the LD50 for PLTX. Then, the toxicity of the Ostreopsis spp. cultures was checked. Several mice were injected with 5 decreased quantities of O. ovata A culture, 139 μM of PLTX-equivalent, O. ovata B culture, 56 μM of PLTX-equivalent, and O. siamensis culture 12.72 μM of PLTXequivalent. From these experiments, 46.5 μg of PLTX equivalent/kg mice weight, 187.5 μg of PLTX equivalent/kg mice weight and 136.5 μg of PLTX equivalent/kg mice weight respectively were obtained as LD50 for the Ostreopsis spp. cultures indicating a higher toxicity in the case of the O. ovata A culture.
| Extract | Percentage of death mouse (%) | LD50 (μg/kg) | Symptoms | ||
|---|---|---|---|---|---|
| 0% | 50% | 100% | |||
| PLTX standard | 0.5 μg/kg | 0.75 μg/kg | 1 μg/kg | 0.75 | Ataxia, spams, dyspnea, scruffy |
| O. ovata A | 18.6 μg/kg | 46.5 μg/kg | 93 μg/kg | 46.5 | |
| O. ovata B | 112.5 μg/kg | 187.5 μg/kg | 375 μg/kg | 187.5 | |
| O. siamensis | 68 μg/kg | 136.5 μg/kg | 170.5 μg/kg | 136.5 | |
| Methanol control | 0.50% | ----- | ----- | ----- | No symptoms observed |
| 0.25% | ----- | ----- | ----- | ||
| 0.125% | ----- | ----- | ----- | ||
| Matrix effect (Coolia monotis) | 0.50% | ----- | ----- | ----- | |
| 0.25% | ----- | ----- | ----- | ||
Table 2: Mouse bioassay data (PLTX-equivalents) after i.p. injection of O. ovata A, O. ovata B strains and O. siamensis. Injections were made diluted in 1 mL of saline solution with 1% Tween 60. A range of 5 concentrations, 0.05-1 μg/Kg, of PLTX standard were used. The mice were observed the first hour and monitored for 24 hours and death mice were registered in addition with symptoms. One lower concentration than those that produce 0% of mouse death and one higher concentration than those that produce 100% of mouse death were also tested in Ostreopsis strains but only three mice were used in each case with the same result than previous concentration. Controls with different methanol percentage and matrix effect were done (three mice per condition) and no effects were observed.
The production of toxins in vitro plays an important role to obtain standards to develop toxicological and pharmacological studies and more accurate detection methods. More important, the knowledge of the optimal conditions for these species to grow better and produce more toxin help us predict where and when their blooms can suddenly appear. As several studies demonstrate, Ostreopsis spp. are changing their world distribution [20-23] and therefore it is necessary to know the strains and species arriving, as well as their toxin profile in order to clarify if any environmental variation can affect their presence.
It was described that only the Japanese O. siamensis produces ostreocin [34], however other studies demonstrate that also Mediterranean O. siamensis synthesizes this toxin [13,35,36]. In addition PLTX or putative PLTX (depending on the author) have been identified in this specie [4,37,38]. Although the term PLTX-like compound is used to designate toxins coming from Ostreopsis spp [13]. This controversy could be due to the lack of standards that prevent the clarification of the toxins produced by each strain. In this sense, many times the results are shown as PLTX-like compounds because although several analogs have been identified, as far as we know, certified material or standards for these products are not yet available. Therefore the identification studies published are based on theoretical chemical approaches from a PLTX standard [5,7-10,18].
The first observation from these results is the fact that to quantify the growth of cultures, the pellet weight is as accurate as the cells number. Since Ostreopsis spp. are benthic dinoflagellate that grow in lumps and segregate a mucous substance the homogenization of the suspension is really hard and some chemical disaggregation is necessary [25,31]. If only the number of cells is measured, the chemical disaggregation with HCl is a useful tool, but when the cultures integrity is important the acidification could be a problem. Although the weight is increased due to the mucous presence, this increase should be proportional with the cells number. In any case, the way these cells grow up and the counting procedure is important since these species are regularly monitored in the water column in some countries to avoid human intoxications. In this sense, a large number of cells could be present in the water but the cells number could be underestimated since most of them are growing in the rock surface or in the seabed making lumps. Therefore, it is very important to bear in mind these data to develop a representative sampling procedure in order to avoid an underestimation in the cell number.
O. ovata produces more PLTX-like compounds than O. siamensis. It is known that these two species produce different PLTX analoges, O. ovata produces ovatoxin-a [36,39] while ostreocin-d is synthesized by O. siamensis [34,40]. Probably these two PLTX-like compounds have different affinity for the Na,K-ATPase. From our data, it seems that ostreocin-d, or the analog produced by this O. siamensis strain, could show less bind to the pump, providing a lower FP signal, and therefore lower PLTX-like concentration. But attending to the growth at 19ºC and 24º C, it seems that in any case, O. siamensis growth is lower than O. ovata, therefore maybe the toxin synthesis is also lower.
Different approaches have been published about the optimal temperature of Ostreopsis blooms occurrence. A temperature of 23°C has been proposed as the optimal temperature to these cells after a bloom in the Adriatic Sea [19]. On the other hand, high abundance of O. ovata has been described when the seawater temperature was 26- 30°C in The Thyrrhenian Sea and in the Genoa coast, while 28-29°C were registered along the Italian coastline of Lazio in 1994 during an Ostreopsis bloom [26,41,42]. Recently, in Japan´s coast a temperature range between 22-30°C depending on the Ostreopsis spp. strain studied has been suggested [43]. Under controlled conditions high growth was observed at 20°C although higher toxin production was obtained at 25°C [31], the same as Ostreopsis lenticularis that produces more toxin at highest temperatures [44]. In addition, it is not clear the influence of the salinity in the toxin production [26,31]. Our results suggest that the toxin production and the growth are higher at 37‰ of salinity and 24º C. This is in accordance with some blooms of Ostreopsis spp. in the Mediterranean Sea when the salinity was around 37-38‰ [22,42]. However 33‰ was described as the optimal salinity [45]. In any case, the same species or strains can grow in different ecological conditions.
Furthermore, the lethal dose of the cultures was checked by bioassay, to know the toxicity of PLTX-like compounds produced by Ostreopsis spp. cultures. The European Food Safety Authority (EFSA) establishes the LD50 after i.p. injections of PLTX around 150-720 ng/ Kg of mice body weight [46]. Our positive PLTX standard control was lethal to the mice at the concentration of 0.750 μg/kg. Ostreopsis spp. mousse bioassay data show that the cultures can be between 40 and 200 times less toxic than the PLTX standard indicating that other toxin with low toxicity and probably lower Na,K-ATPase affinity than PLTX is present in the cultures. From our results, O. ovata A strain is more toxic than O. ovata B and O. siamensis, since around 4 times more extract is necessary to induce the mice death.
In addition, different toxicity as well as different results after FP detection were shown by both O. ovata strains. Therefore, depending on the strain and the culture conditions, a larger amount or more toxic compounds can be produced. Due to the lack of standards, it is very difficult to establish the toxin profiles of the samples. By using different cultures and according to the toxin profile identified in each, the range of toxin potency proposed in the bibliography was PLTX>ostreocind> mascarenotoxin -a, and ovatoxin-a was not classified [13,47]. Taking into account our results and based on this classification, if ostreocin-d is present in O. siamensis, the compound produced by O. ovata A, probably ovatoxin-a, is more toxic and shows higher affinity by the Na,K-ATPase as FP results shows. However several analogs could be present in each strain and probably depending on the conditions maybe one can be prevalent. Therefore the identification of compounds as well as the equivalence of bioassay and Na,K-ATPase affinity should be a further approach when standards become available in order to monitor and predict the presence of toxic strains in temperate regions.
In summary, O. ovata produces more PLTX-like compounds than O. siamensis. A salinity of 37‰, 24°C, and 16:8 h light-dark photoperiod are the optimal conditions to grow Ostreopsis spp. cultures and produce PLTX-like compounds. In addition, another relevant conclusion of this work is that sampling of the algae for a monitoring program can not be done directly on the water given their growth habitat.
Ostreopsis ovata strains were kindly provided by Dra. K. Aligizaki from Thessaloniki University, Greece. Ostreopsis siamensis cultures were provided by Laboratorio de Recursos Pesqueros, Huelva (Spain). The authors wish to thank Dra. K. Aligizaki from Thessaloniki University, Greece for her kind gift of some of the strains used. This work was funded with the following FEDER cofundedgrants: From Ministerio de Ciencia y Tecnología, Spain: AGL2009-13581- CO2-01, AGL2012-40485-CO2-01. From Xunta de Galicia, Spain: 10PXIB261254 PR. From EU VIIth Frame Program: 211326–CP (CONffIDENCE), 265896 BAMMBO, 265409 μA QUA, and 262649 BEADS, 315285 Ciguatools and 312184 PharmaSea. From the European Union’s Seventh Framework Programme managed by REA–Research Executive Agency http://ec.europa.eu/research/ rea (FP7/2007-2013) under grant agreement Nos. 211326 – CP (CONffIDENCE), 265896 BAMMBO, 265409 μAQUA, and 262649 BEADS, 315285 Ciguatools and 312184 PharmaSea. From the Atlantic Area Programme (Interreg IVB Transnational): 2009-1/117 Pharmatlantic. Andrea Fernández Araujo is supported by a fellowship from Subprograma de Formación de Personal Investigador (AGL2009- 13581-C02-01), Spain.